
IT’S ALL IN THE GENES
As dog breeders, we engage in genetic “experiments” each time we plan a mating. The type of mating selected should coincide with your goals. To some breeders, determining which traits will appear in the offspring of a mating is like rolling the dice – a combination of luck and chance. For others, producing certain traits involves more skill than luck – the result of careful study and planning. As breeders, we must understand how we manipulate genes within our breeding stock to produce the kinds of dogs we want. We have to first understand dogs as a species, then dogs as genetic individuals.
The species, Canis familiaris, includes all breeds of the domestic dog. Although we can argue that there is little similarity between a Chihuahua and a Saint Bernard, or that established breeds are separate entities among themselves, they all are genetically the same species. While a mating within a breed may be considered outbred, it still must be viewed as part of the whole genetic picture: a mating within an isolated, closely related, interbred population. Each breed was developed by close breeding and inbreeding among a small group of founding canine ancestors, either through a long period of genetic selection or by intensely inbreeding a smaller number of generations. The process established the breed’s characteristics and made the dogs in it breed true.
When evaluating your breeding program, remember that most traits you’re seeking cannot be changed, fixed or created in a single generation. The more information you can obtain on how certain traits have been transmitted by your dog’s ancestors, the better you can prioritize your breeding goals. Tens of thousands of genes interact to produce a single dog. All genes are inherited in pairs, one pair from the father and one from the mother. If the pair of inherited genes from both parents is identical, the pair is called homozygous. If the genes in the pair are not alike, the pair is called heterozygous. Fortunately, the gene pairs that make a dog a dog and not a cat are always homozygous. Similarly, the gene pairs that make a certain breed always breed true are also homozygous. . Therefore, a large proportion of homozygous non-variable pairs – those that give a breed its specific standard – exist within each breed. It is the variable gene pairs, like those that control color, size and angulation, that produce variations within a breed.
BREEDING BY PEDIGREE
Outbreeding brings together two dogs less related than the average for the breed. This promotes more heterozygosity, and gene diversity within each dog by matching pairs of unrelated genes from different ancestors. Outbreeding can also mask the expression of recessive genes, and allow their propagation in the carrier state.
Most outbreeding tends to produce more variation within a litter. An exception would be if the parents are so dissimilar that they create a uniformity of heterozygosity. This is what usually occurs in a mismating between two breeds. The resultant litter tends to be uniform, but demonstrates “half-way points” between the dissimilar traits of the parents. Such litters may be phenotypically uniform, but will rarely breed true due to the mix of dissimilar genes.
A reason to outbreed would be to bring in new traits that your breeding stock does not possess. While the parents may be genetically dissimilar, you should choose a mate that corrects your dog’s faults but phenotypically complements your dog’s good traits.

It is not unusual to produce an excellent quality dog from an outbred litter. The abundance of genetic variability can place all the right pieces in one individual. Many top-winning show dogs are outbred. Consequently, however, they may have low inbreeding coefficients and may lack the ability to uniformly pass on their good traits to their offspring. After an outbreeding, breeders may want to breed back to dogs related to their original stock, to increase homozygosity and attempt to solidify newly acquired traits.
Linebreeding attempts to concentrate the genes of a specific ancestor or ancestors through their appearance multiple times in a pedigree. The ancestor should appear behind more than one offspring. If an ancestor always appears behind the same offspring, you are only linebreeding on the approximately 50 percent of the genes passed to the offspring and not the ancestor itself.
It is better for linebred ancestors to appear on both the sire’s and the dam’s sides of the pedigree. That way their genes have a better chance of pairing back up in the resultant pups. Genes from common ancestors have a greater chance of expression when paired with each other than when paired with genes from other individuals, which may mask or alter their effects.
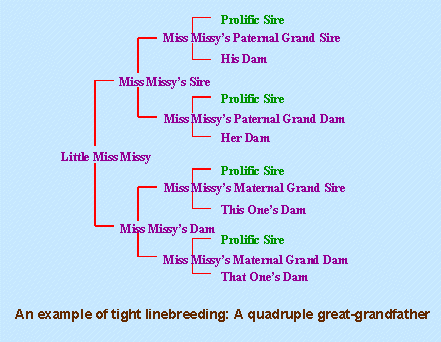
A linebreeding may produce a puppy with magnificent qualities, but if those qualities are not present in any of the ancestors the pup has been linebred on, it may not breed true. Therefore, careful selection of mates is important, but careful selection of puppies from the resultant litter is also important to fulfill your genetic goals. Without this, you are reducing your chances of concentrating the genes of the linebred ancestor.
Increasing an individual’s homozygosity through linebreeding may not, however, reproduce an outbred ancestor. If an ancestor is outbred and generally heterozygous (Aa), increasing homozygosity will produce more AA and aa. The way to reproduce an outbred ancestor is to mate two individuals that mimic the appearance and pedigree of the ancestor’s parents.
Inbreeding significantly increases homozygosity, and therefore uniformity in litters. Inbreeding can increase the expression of both beneficial and detrimental recessive genes through pairing up. If a recessive gene (a) is rare in the population, it will almost always be masked by a dominant gene (A). Through inbreeding, a rare recessive gene (a) can be passed from a heterozygous (Aa) common ancestor through both the sire and dam, creating a homozygous recessive (aa) offspring. Inbreeding does not create undesirable genes, it simply increases the expression of those that are already present in a heterozygous state.

Inbreeding can exacerbate a tendency toward disorders controlled by multiple genes, such as hip dysplasia and congenital heart anomalies. Unless you have prior knowledge of what milder linebreedings on the common ancestors have produced, inbreeding may expose your puppies (and puppy buyers) to extraordinary risk of genetic defects. Research has shown that inbreeding depression, or diminished health and viability through inbreeding is directly related to the amount of detrimental recessive genes present. Some lines thrive with inbreeding, and some do not.
PEDIGREE ANALYSIS
Geneticists’ and breeders’ definitions of inbreeding vary. A geneticist views inbreeding as a measurable number that goes up whenever there is a common ancestor between the sire’s and dam’s sides of the pedigree; a breeder considers inbreeding to be close inbreeding, such as father-to-daughter or brother-to-sister matings. A common ancestor, even in the eighth generation, will increase the measurable amount of inbreeding in the pedigree.
The Inbreeding Coefficient (or Wright’s coefficient) is an estimate of the percentage of all the variable gene pairs that are homozygous due to inheritance from common ancestors. It is also the average chance that any single gene pair is homozygous due to inheritance from a common ancestor. In order to determine whether a particular mating is an outbreeding or inbreeding relative to your breed, you must determine the breed’s average inbreeding coefficient. The average inbreeding coefficient of a breed will vary depending on the breed’s popularity or the age of its breeding population. A mating with an inbreeding coefficient of 14 percent based on a ten generation pedigree, would be considered moderate inbreeding for a Labrador Retriever (a popular breed with a low average inbreeding coefficient), but would be considered outbred for an Irish Water Spaniel (a rare breed with a higher average inbreeding coefficient).
For the calculated inbreeding coefficient of a pedigree to be accurate, it must be based on several generations. Inbreeding in the fifth and later generations (background inbreeding) often has a profound effect on the genetic makeup of the offspring represented by the pedigree. In studies conducted on dog breeds, the difference in inbreeding coefficients based on four versus eight generation pedigrees varied immensely. A four generation pedigree containing 28 unique ancestors for 30 positions in the pedigree could generate a low inbreeding coefficient, while eight generations of the same pedigree, which contained 212 unique ancestors out of 510 possible positions, had a considerably higher inbreeding coefficient. What seemed like an outbred mix of genes in a couple of generations, appeared as a linebred concentration of genes from influential ancestors in extended generations.
The process of calculating coefficients is too complex to present here. Several books that include how to compute coefficients are indicated at the end of this article; some computerized canine pedigree programs also compute coefficients. The analyses in this article were performed using CompuPed, by RCI Software.
[RCI Note: CompuPed computes Wright’s Inbreeding Coefficient faster and more accurately than any other PC program available. ]
Pedigree of: “Laurel Hill Braxfield Bilye”
( a spayed female Gordon Setter owned by Dr. Jerold and Mrs. Candice Bell, and co-bred by Mary Poos and Laura Bedford.)
1 2 3 4 5
Dual CH Loch Adair Monarch
CH Sutherland MacDuff
| CH Sutherland Dunnideer Waltz
CH Sutherland Gallant
| | CH Afternod Kyle of Sutherland
| CH Sutherland Pavane
| CH Sutherland Xenia
CH Loch Adair Foxfire
| | Afternod Fidemac
| | CH Loch Adair Peer of Sutherland, CD
| | CH Wee Laurie Adair
| CH Sutherland Lass of Shambray
| | CH Afternod Callant
| CH Afternod Karma
| CH Afternod Amber
CH Braxfield Andrew of Aberdeen
| | Afternod Fidemac
| | AmCnCH Afternod Scot of Blackbay, CD
| | | CH Afternod Alder
| | AmCnCH Forecast Trade Winds, CD
| | | | Bud O'Field Brookview
| | | CH Oak Lynn's Bonnie Bridget
| | | Borderland Taupie
| CH Afternod Ember VI, CD
| | CH Afternod Simon
| | Afternod Profile of Sark
| | | CH Afternod Heiress of Sark
| CH Afternod Ember V
| | CH Afternod Callant
| CH Afternod Maud MacKenzie
| CH Afternod Amber
LAUREL HILL BRAXFIELD BILYE
| CH Afternod Callant
| Dual CH Loch Adair Monarch
| | Loch Adair Diana of Redchico
| CH Sutherland MacDuff
| | | CH Afternod Anagram
| | CH Sutherland Dunnideer Waltz
| | CH Hi-Laway's Calopin
| CH Kendelee Pendragon
| | | CH Afternod Callant
| | | CH Wee Jock Adair, CD
| | | | Loch Adair Diana of Redchico
| | CH Afternod Nighean Kendelee
| | | CH Afternod Simon
| | CH Afternod Wendee
| | Afternod Dee of Aberdeen
CH Halcyon Belle-Amie
| Dual CH Loch Adair Monarch
| CH Sutherland MacDuff
| | CH Sutherland Dunnideer Waltz
| CH Sutherland Gallant
| | | CH Afternod Kyle of Sutherland
| | CH Sutherland Pavane
| | CH Sutherland Xenia
CH Loch Adair Firefly, WD
| Afternod Fidemac
| CH Loch Adair Peer of Sutherland, CD
| | CH Wee Laurie Adair
CH Sutherland Lass of Shambray
| CH Afternod Callant
CH Afternod Karma
CH Afternod Amber
To visualize some of these concepts, please refer to the above pedigree. Linebred ancestors in this pedigree are in color, to help visualize their contribution. The paternal grandsire, CH Loch Adair Foxfire, and the maternal grandam, CH Loch Adair Firefly WD, are full siblings, making this a first-cousin mating. The inbreeding coefficient for a first cousin mating is 6.25%, which is considered a mild level of inbreeding. Lists of inbreeding coefficients based on different types of matings are shown in the table below.

In Bilye’s pedigree, an inbreeding coefficient based on four generations computes to 7.81%. This is not significantly different from the estimate based on the first-cousin mating alone. Inbreeding coefficients based on increasing numbers of generations are as follows: five generations, 13.34%; six generations, 18.19%; seven generations, 22.78%; eight generations, 24.01%; ten generations, 28.63%; and twelve generations, 30.81%. The inbreeding coefficient of 30.81 percent is more than what you would find in a parent-to-offspring mating (25%). As you can see, the background inbreeding has far more influence on the total inbreeding coefficient than the first-cousin mating, which only appears to be its strongest influence.
Knowledge of the degree of inbreeding in a pedigree does not necessarily help you unless you know whose genes are being concentrated. The percent blood coefficient measures the relatedness between an ancestor and the individual represented by the pedigree. It estimates the probable percentage of genes passed down from a common ancestor. We know that a parent passes on an average of 50% of its genes, while a grandparent passes on 25%, a great-grandparent 12.5%, and so on. For every time the ancestor appears in the pedigree, its percentage of passed-on genes can be added up and its “percentage of blood” estimated.
In many breeds, an influential individual may not appear until later generations, but then will appear so many times that it necessarily contributes a large proportion of genes to the pedigree. This can occur in breeds, due to either prolific ancestors (usually stud dogs), or with a small population of dogs originating the breed. Based on a twenty-five generation pedigree of Bilye, there are only 852 unique ancestors who appear a total of over twenty-million times.
Pedigree Analysis of Laurel Hill Braxfield Bilye
(computed to 25 generations)
1st Generation |
|||
Linebred Ancesters |
Percentage of blood |
Appearance in pedigree |
# times in pedigree |
| CH Afternod Drambuie | 33.20% | 6 | 33 |
| CH Afternod Sue | 27.05% | 7 | 61 |
| CH Afternod Callant | 26.56% | 5 | 13 |
| “Grand-Parents” | 25.00% | 2 | 1 |
| CH Sutherland Gallant | 25.00% | 3 | 2 |
| CH Sutherland MacDuff | 25.00% | 3 | 3 |
| CH Sutherland Lass of Shambray | 25.00% | 3 | 2 |
| CH Wilson’s Corrie, CD | 22.30% | 7 | 200 |
| CH Afternod Buchanon | 20.22% | 7 | 48 |
| Loch Adair Diana of Redchic | 17.97% | 5 | 12 |
| CH EEG’s Scotia Nodrog Rettes | 17.76% | 8 | 181 |
| Afternod Ember of Gordon Hill | 17.14% | 8 | 76 |
| CH Afternod Hickory | 16.21% | 6 | 27 |
| CH Black Rogue of Serlway | 15.72% | 9 | 480 |
| CH Afternod Woodbine | 14.45% | 6 | 15 |
| CH Fast’s Falcon of Windy Hill | 13.82% | 8 | 66 |
| Afternod Fidemac | 13.67% | 5 | 7 |
| CH Page’s MacDonegal II | 13.43% | 7 | 56 |
| Afternod Hedera | 13.38% | 7 | 56 |
| CH Downside Bonnie of Serlway | 12.90% | 10 | 708 |
| Peter of Crombie | 12.76% | 11 | 3,887 |
| “Great-Grand-Parents” | 12.50% | 3 | 1 |
| CH Afternod Amber | 12.50% | 5 | 5 |
| Ben of Crombie | 11.83% | 11 | 7,584 |
| Stylish William | 11.18% | 13 | 23,764 |
| Stylish Billie | 11.08% | 14 | 70,542 |
| Stylish Ranger | 10.80% | 15 | 297,331 |
| CH Afternod Kate | 10.74% | 6 | 17 |
| Heather Grouse | 10.61% | 16 | 1,129,656 |
| Afternod Hedemac | 10.45% | 7 | 28 |
The above analysis shows the ancestral contribution of the linebred ancestors in Bilye’s pedigree. Those dogs in color were present in the five-generation pedigree. CH Afternod Drambuie has the highest genetic contribution of all of the linebred ancestors. He appears 33 times between the sixth and eighth generations. One appearance in the sixth generation contributes 1.56% of the genes to the pedigree. His total contribution is 33.2% of Bilye’s genes, second only to the parents. Therefore, in this pedigree, the most influential ancestor doesn’t even appear in the five-generation pedigree. His dam, CH Afternod Sue, appears 61 times between the seventh and tenth generations, and contributes more genes to the pedigree than a grandparent.
Foundation dogs that formed the Gordon Setter breed also play a great role in the genetic makeup of today’s dogs. Heather Grouse appears over one million times between the sixteenth and twenty-fifth generations, and almost doubles those appearances beyond the twenty-fifth generation. He contributes over ten percent of the genes to Bilye’s pedigree. This example shows that the depth of the pedigree is very important in estimating the genetic makeup of an individual. Any detrimental recessive genes carried by Heather Grouse or other founding dogs, would be expected to be widespread in the breed.
BREEDING BY APPEARANCE
Many breeders plan matings solely on the appearance of a dog and not on its pedigree or the relatedness of the prospective parents. This is called assortative mating. Breeders use positive assortative matings (like-to-like) to solidify traits, and negative assortative matings (like-to-unlike) when they wish to correct traits or bring in traits their breeding stock may lack.
Some individuals may share desirable characteristics, but they inherit them differently. This is especially true of polygenic traits, such as ear set, bite, or length of forearm. Breeding two phenotypically similar but genotypically unrelated dogs together would not necessarily reproduce these traits. Conversely, each individual with the same pedigree will not necessarily look or breed alike.
Breedings should not be planned solely on the basis of the pedigree or appearance alone. Matings should be based on a combination of appearance and ancestry. If you are trying to solidify a certain trait – like topline – and it is one you can observe in the parents and the linebred ancestors of two related dogs, then you can be more confident that you will attain your goal.
GENETIC DIVERSITY
Some breed clubs advocate codes of ethics that discourage linebreeding or inbreeding, as an attempt to increase breed genetic diversity. This position is based on a falsle premise. Inbreeding or linebreeding does not cause the loss of genes from a breed gene pool. It occurs through selection; the use and non-use of offspring. If some breeders linebreed to certain dogs that they favor, and others linebreed to other dogs that they favor, then breed-wide genetic diversity is maintained.
In a theoretical mating with four offspring, we are dealing with four gene pairs. The sire is homozygous at 50% of his gene pairs (two out of four), while the dam is homozygous at 75% of her gene pairs. It is reasonable to assume that she is more inbred than the sire.
A basic tenet of population genetics is that gene frequencies do not change from the parental generation to the offspring. This will occur regardless of the homozygosity or heterozygosity of the parents, or whether the mating is an outbreeding, linebreeding, or inbreeding. This is the nature of genetic recombination.

There is a lack of gene diversity at the first (olive) gene pair, so that only one type of gene combination can be produced: homozygous olive. As the sire is homozygous lime at the third gene pair, and the dam is homozygous blue, all offspring will be heterozygous at the third gene pair. Depending on the dominant or recessive nature of the blue or lime genes, all offspring will appear the same for this trait due to a uniformity of heterozygosity.
If offspring D is used as a prolific breeder, and none of the other offspring are bred to a great extent, gene frequencies in the breed will change. As dog D lacks the orange gene in the second pair and the purple gene in the fourth pair, the frequencies of these genes will diminish in the breed. They will be replaced by higher frequencies of the red and pink genes. This shifts the gene pool, and the breed’s genetic diversity. Of course, dogs have more than four gene pairs, and the overuse of dog D to the exception of others can affect the gene frequency of thousands of genes. Again, it is selection (for example of dog D to the exception of others), and not the types of matings he is involved in that alters gene frequencies.
Breeders should select the best individuals from all kennel lines, so as to not create new genetic bottlenecks. There is a tendency for many breeders to breed to a male; who produced no epileptics in matings to several epileptic dams, to an OFA excellent stud, or to the top winning dog in the show ring. Regardless of the popularity of the breed, if everyone is breeding to a single studdog, (the popular sire syndrome) the gene pool will drift in that dog’s direction and there will be a loss of genetic diversity. Too much breeding to one dog will give the gene pool an extraordinary dose of his genes, and also whatever detrimental recessives he may carry, to be uncovered in later generations. This can cause future breed related genetic disease through the founders effect.
Dogs who are poor examples of the breed should not be used simply to maintain diversity. Related dogs with desirable qualities will maintain diversity, and improve the breed. Breeders should concentrate on selecting toward a breed standard, based on the ideal temperament, performance, and conformation, and should select against the significant breed related health issues. Using progeny and sib-based information to select against both polygenic disorders and those without a known mode of inheritance will allow greater control.
Rare breeds with small gene pools have concerns about genetic diversity. What constitutes acceptable diversity versus too restricted diversity? The problems with genetic diversity in purebred populations concern the fixing of deleterious recessive genes, which when homozygous cause impaired health. Lethal recessives place a drain on the gene pool either prenatally, or before reproductive age. They can manifest themselves through smaller litter size, or neonatal death. Other deleterious recessives cause disease, while not affecting reproduction.
Problems with a lack of genetic diversity arise at the gene locus level. There is no specific level or percentage of inbreeding that causes impaired health or vigor. It has been shown that some inbred strains of animals thrive generation after generation, while others fail to thrive. If there is no diversity (non-variable gene pairs for a breed) but the homozygote is not detrimental, there is no effect on breed health. The characteristics that make a breed reproduce true to its standard are based on non-variable gene pairs. A genetic health problem arises for a breed when a detrimental allele increases in frequency and homozygosity.
GENETIC CONSERVATION
The perceived problem of a limited gene pool has caused some breeds to advocate outbreeding of all dogs. Studies in genetic conservation and rare breeds have shown that this practice actually contributes to the loss of genetic diversity. By uniformly crossing all “lines” in a breed, you eliminate the differences between them, and therefore the diversity between individuals. This practice in livestock breeding has significantly reduced diversity, and caused the loss of unique rare breeds. The process of maintaining healthy “lines” or families of dogs, with many breeders crossing between lines and breeding back as they see fit maintains diversity in the gene pool. It is the varied opinion of breeders as to what constitutes the ideal dog, and their selection of breeding stock that maintains breed diversity.
The Doberman Pincher breed is large, and genetically diverse. The breed has a problem with vonWillibrands disease, an autosomal recessive bleeding disorder. Some researchers estimate that up to 60% of the breed may be homozygous recessive for the defective gene, and the majority of the remaining dogs are heterozygous. Therefore, there is diminished genetic diversity in this breed at the vonWillibrands locus. A genetic test and screening program now exists for Doberman Pincher breeders. They can identify carrier and affected dogs, and decrease the defective gene frequency through selection of normal testing offspring for breeding. By not just eliminating carriers, but replacing them with normal testing offspring, genetic diversity will be conserved.
Dalmatians have a high frequency defective autosomal recessive gene controlling purine metabolism. Homozygous recessive individuals can have urinary problems due to urate bladder stones and crystals, and an associated skin condition (Dalmatian Bronzing Syndrome). At one time, the breed and the AKC approved a crossbreeding program to a few Pointers, to bring normal purine metabolism genes into the gene pool. The program was abandoned for several reasons, but it was accepted that the number of individual Dalmatians with two normal purine metabolism genes far exceeded the few Pointers that were being used in the program. The impact of other Pointer genes foreign to the Dalmatian gene pool could have had a greater detrimental effect than the few normal purine metabolism genes being imported through the program.
PUTTING IT ALL TOGETHER
Decisions to linebreed, inbreed or outbreed should be made based on the knowledge of an individual dog’s traits and those of its ancestors. Inbreeding will quickly identify the good and bad recessive genes the parents share in the offspring. Unless you have prior knowledge of what the pups of milder linebreedings on the common ancestors were like, you may be exposing your puppies (and puppy buyers) to extraordinary risk of genetic defects. In your matings, the inbreeding coefficient should only increase because you are specifically linebreeding (increasing the percentage of blood) to selected ancestors.
Don’t set too many goals in each generation, or your selective pressure for each goal will necessarily become weaker. Genetically complex or dominant traits should be addressed early in a long-range breeding plan, as they may take several generations to fix. Traits with major dominant genes become fixed more slowly, as the heterozygous (Aa) individuals in a breed will not be readily differentiated from the homozygous-dominant (AA) individuals. Desirable recessive traits can be fixed in one generation because individuals that show such characteristics are homozygous for the recessive genes. Dogs that breed true for numerous matings and generations should be preferentially selected for breeding stock. This prepotency is due to homozygosity of dominant (AA) and recessive (aa) genes.
If you linebreed and are not happy with what you have produced, breeding to a less related line immediately creates an outbred line and brings in new traits. Repeated outbreeding to attempt to dilute detrimental recessive genes is not a desirable method of genetic disease control. Recessive genes cannot be diluted; they are either present or not. Outbreeding carriers multiplies and further spreads the defective gene(s) in the gene pool. If a dog is a known carrier or has high carrier risk through pedigree analysis, it can be retired from breeding, and replaced with one or two quality offspring. Those offspring should be bred, and replaced with quality offspring of their own, with the hope of losing the defective gene.
Trying to develop your breeding program scientifically can be an arduous, but rewarding, endeavor. By taking the time to understand the types of breeding schemes available, you can concentrate on your goals towards producing a better dog.
Further Reading:
If you are interested in learning more about these subjects, consult the following books:
- Abnormalities of Companion Animals: Analysis of Heritability
C.W. Foley, J.F. Lasley, and G.D. Osweiler, Iowa State University Press, Ames, Iowa. 1979. - Genetics for Dog Breeders
F.B. Hutt, W.H. Freeman Co, San Francisco, California. 1979. - Veterinary Genetics
F. W. Nicholas, Clarendon Press, Oxford England. 1987. - Genetics for Dog Breeders
R. Robinson, Pergamon Press, Oxford England. 1990. - Genetics of the Dog (equally applicable to cats & other animals)
M.B. Willis, Howell Book House, New York, New York. 1989.